de vuelta al
capullo
Los recuerdos suelen ocupar una gran porción de las distracciones mentales diarias de una persona corriente. Podemos recordar una comilona de órdago, los juegos a los que jugábamos de niños o una anécdota especialmente hilarante ocurrida en la oficina el día anterior, evocar una puesta de sol sobrecogedora que contemplamos una vez en una playa o rememorar experiencias particularmente traumáticas o aterradoras. Nuestra memoria depende de las percepciones sensoriales: un olor familiar o una canción preferida pueden desencadenar una oleada de recuerdos vívidos que nos retrotraen a un momento o un lugar concretos.
Como hemos visto, las plantas también tienen percepciones sensoriales muy diversas. Sin embargo, es evidente que carecen de recuerdos comparables a los nuestros: no se encogen de miedo al pensar en una sequía ni sueñan con los rayos de sol del verano. Tampoco echan de menos estar encerradas en una vaina ni se ponen nerviosas con la liberación prematura del polen. A diferencia de la Abuela Sauce de la película de Disney Pocahontas, los árboles vetustos no recuerdan la historia de las personas que han dormido a su sombra. Ahora bien, tal como hemos visto en capítulos anteriores, es evidente que tienen la capacidad de retener eventos pretéritos y de recuperar esa información en un momento posterior para integrarla en su marco de desarrollo: las plantas del tabaco saben el color de la última luz que han visto y los sauces saben si las orugas han atacado a sus vecinos. Estos ejemplos, entre muchos otros, ilustran una reacción retardada a un suceso previo, un elemento clave de la memoria.
Mark Jaffe, el mismo científico que acuñó el término de «tigmomorfogénesis», publicó uno de los primeros estudios sobre la memoria de las plantas en 1977, si bien no hablaba de ella en tales términos, sino que se refería a una retención de entre una y dos horas de duración de la información sensorial asimilada.[1] A Jaffe le interesaba averiguar qué induce a los zarcillos del guisante a curvarse cuando tocan un objeto al que pueden enroscarse. Los zarcillos del guisante son estructuras con forma de tallo que crecen en línea recta hasta que tropiezan con una valla o un poste que pueden utilizar como apoyo, momento a partir del cual se enroscan rápidamente al objeto para aferrarse a él.
Jaffe demostró que si cortaba un zarcillo de una planta del guisante pero lo mantenía en un entorno húmedo y bien iluminado, podía conseguir que el zarcillo amputado se enroscase simplemente frotándole la cara inferior con un dedo. Sin embargo, cuando llevó a cabo el mismo experimento en la oscuridad, los zarcillos extirpados no se enrollaban cuando los tocaba, lo cual indicaba que necesitaban luz para realizar sus giros mágicos. Con todo, el dato verdaderamente interesante era el siguiente: si un zarcillo al cual se había tocado en la oscuridad se colocaba a la luz una o dos horas después, se enroscaba de manera espontánea, sin necesidad de que Jaffe lo frotara de nuevo. De algún modo, observó Jaffe, el zarcillo que había tocado en la oscuridad había retenido esa información y la había recuperado una vez depositado bajo la luz. ¿Podía considerarse este almacenamiento y posterior recuperación de información una especie de «memoria»?
Las investigaciones en materia de memoria humana llevadas a cabo por el reputado psicólogo Endel Tulving nos proporcionan un fundamento inicial a partir del cual explorar las plantas y sus «recuerdos» únicos. Tulving propuso que la memoria humana existe en tres niveles.[2] El nivel inferior, la memoria procedimental, alude a un recuerdo no verbal de cómo hacer las cosas y depende de la capacidad de percibir estímulos externos (como recordar nadar al saltar a una piscina). En el segundo nivel se encuentra la memoria semántica, la memoria de conceptos (que engloba, por ejemplo, la mayoría de las materias que aprendimos en la escuela). Y el tercer nivel es la memoria episódica, que consiste en recordar eventos autobiográficos, como disfraces divertidos que nos pusimos en fiestas de Halloween en nuestra infancia o la tristeza que sentimos tras la muerte de una mascota a la que teníamos mucho cariño. La memoria episódica depende de la «conciencia propia» de cada persona. Es evidente que las plantas no dan la talla para la memoria semántica o episódica, que en realidad son las memorias que nos definen como seres humanos. Sin embargo, sí son capaces de notar estímulos externos y reaccionar, de manera que, de acuerdo con la definición de Tulving, las plantas tendrían memoria procedimental.[3] Y en efecto, así lo ilustran las plantas de guisantes de Jaffe, que notaban su tacto, lo recordaban y reaccionaban enroscándose.
Los neurobiólogos saben bastante de fisiología de la memoria y son capaces de identificar las zonas diferenciadas (más interconectadas) del cerebro responsables de los distintos tipos de memoria. Los científicos saben que es imprescindible que existan señales eléctricas entre las neuronas para que se formen y archiven recuerdos. En cambio, sabemos mucho menos acerca de la base molecular y celular de la memoria. Lo más fascinante es que las últimas investigaciones apuntan a que, si bien los recuerdos son infinitos, el número de proteínas implicadas en su conservación es reducido.[4]
Huelga decir que lo que denominamos «memoria» en el caso de los humanos en realidad es un término que abarca muchas formas distintas de memoria, además de las descritas por Tulving. Tenemos memoria sensorial, que recibe y filtra rápidamente las percepciones de los sentidos (en un pestañeo); memoria a corto plazo, que permite retener unos siete objetos en la conciencia durante varios segundos, y memoria a largo plazo, que define nuestra capacidad de almacenar recuerdos durante toda la vida. También tenemos memoria muscular, un tipo de memoria procedimental consistente en aprender de manera inconsciente movimientos como los que hacemos con los dedos para atarnos los cordones de los zapatos, y memoria inmunológica, gracias a la cual nuestro sistema inmunológico recuerda infecciones pasadas para evitar otras futuras. Salvo la última, todas ellas dependen de funciones cerebrales. La memoria inmunológica, por su parte, depende del funcionamiento de los glóbulos blancos y los anticuerpos.
Pero todas las formas de memoria tienen en común que incluyen los procesos de formación de recuerdos (codificación de información), retención de los recuerdos (almacenamiento de información) y evocación del recuerdo (recuperación de la información). Incluso la memoria informática utiliza exactamente estos tres procesos. Y si queremos detectar aunque sea la memoria más sencilla en las plantas, estos son los procesos que debemos observar.
LA MEMORIA A CORTO PLAZO DE LA VENUS ATRAPAMOSCAS
Tal como hemos visto en el capítulo 4, la venus atrapamoscas necesita saber cuándo un bocado ideal camina sobre sus hojas. Cerrar su trampa exige una inversión inmensa de energía y volver a abrirla puede llevarle varias horas, de manera que la Dionaea solo acciona el muelle cuando está segura de que el insecto que camina despacito por su superficie es lo bastante grande como para dedicarle esa inversión de tiempo. Los grandes filamentos negros de sus lóbulos permiten a la venus atrapamoscas tocar literalmente a su presa, y son esos pelos los que accionan el muelle que cierra la trampa cuando la presa adecuada se interna en ella. Si el insecto roza solo uno de ellos, la trampa permanece abierta. En cambio, lo más probable es que un insecto lo bastante grande toque dos de esos pelos en un margen de unos veinte segundos, y esa señal hace que la venus atrapamoscas entre en acción.
Este sistema puede considerarse análogo a la memoria a corto plazo. En primer lugar, la venus atrapamoscas codifica la información (forma el recuerdo) de que algo (no sabe qué) ha tocado uno de sus pelos. A continuación, almacena esa información durante unos segundos (retiene el recuerdo) y finalmente la recupera (evoca el recuerdo) cuando le tocan un segundo filamento. Si una hormiguita tarda un rato en desplazarse de un pelo al otro, la trampa habrá olvidado el primer contacto cuando el insecto roce el segundo pelo. Dicho de otro modo, pierde la información almacenada, la trampa no se cierra y la hormiga continúa caminando alegremente por la hoja. ¿Cómo codifica y guarda la planta la información del roce del insecto desprevenido con el primer filamento? ¿Y cómo recuerda ese primer contacto para reaccionar al segundo?
Estos interrogantes fueron un enigma para la ciencia desde que John Burdon-Sanderson publicó el primer informe sobre la fisiología de la venus atrapamoscas, datado en 1882.[5] Un siglo después, Dieter Hodick y Andreas Sievers de la Universidad de Bonn, en Alemania, propusieron que la venus atrapamoscas almacenaba información acerca de cuántos pelos le habían tocado en la carga eléctrica de su hoja.[6] Su modelo resulta elegante por su simplicidad. En sus estudios descubrieron que el contacto con uno de los filamentos accionadores de la venus atrapamoscas provoca un potencial de acción eléctrico que induce la apertura de los canales de calcio en la trampa (esta combinación de los potenciales de acción y la apertura de los canales de calcio es similar a los procesos que tienen lugar durante la comunicación neuronal en los seres humanos) y, por ende, provoca un rápido aumento en la concentración de iones de calcio.
Plantearon asimismo que la trampa requiere una concentración de calcio relativamente elevada para cerrarse y que un único potencial de acción provocado por el contacto con un único pelo inductor no alcanza dicho nivel. En consecuencia, es preciso estimular un segundo filamento para incrementar la concentración de calcio en este umbral y accionar la trampa. La codificación de la información se da en el aumento inicial de los niveles de calcio. Para retener la información es preciso mantener un nivel lo bastante elevado de calcio como para que un segundo incremento (accionado por el contacto con un segundo pelo) sobrepase el umbral de calcio concentrado. Dado que las concentraciones de calcio se disipan con el tiempo, si el segundo contacto y potencial no acontecen enseguida, la concentración final tras el segundo desencadenante no es lo bastante elevada para cerrar la trampa y el recuerdo se pierde.
Estudios de investigación posteriores corroboran este modelo. Alexander Volkov y sus colegas de la Oakwood University, Alabama, fueron los primeros en demostrar que, en efecto, es la electricidad la que hace que la venus atrapamoscas se cierre de golpe.[7] Para comprobar su modelo, instalaron electrodos diminutos y aplicaron una descarga eléctrica a los lóbulos abiertos de la planta. De este modo consiguieron que la trampa se cerrara sin existir un contacto directo con sus filamentos sensibles (y aunque no midieron los niveles de calcio, es probable que la corriente provocara su incremento). A continuación modificaron el experimento alterando la cantidad de corriente eléctrica, cosa que permitió a Volkov determinar la descarga exacta necesaria para que la trampa se cierre: se precisaba un flujo de catorce microculombios (una cantidad ligeramente superior a la de la electricidad estática generada al frotar dos globos entre sí) entre los dos electrodos. Dicha descarga podía producirse en una sola ráfaga o como una serie de descargas más breves en un plazo de veinte segundos. Si la carga total tardaba más de veinte segundos en acumularse, la trampa permanecía abierta.
De manera que este es el mecanismo que explica la memoria a corto plazo de la venus atrapamoscas. El primer contacto con un pelo activa un potencial eléctrico que radia de célula en célula. Esta descarga eléctrica se almacena a modo de incremento en las concentraciones de iones durante un breve lapso, y transcurridos unos veinte minutos se disipa. Pero si un segundo potencial de acción entra en contacto con la nervadura central en este periodo, la carga acumulativa y las concentraciones de iones superan el umbral y la trampa se cierra. Si transcurre demasiado tiempo entre ambos potenciales de acción, la planta olvida el primero y la trampa permanece abierta.
Esta señal eléctrica en la venus atrapamoscas (y, por extensión, las señales eléctricas en otras plantas) es similar a las señales eléctricas que intercambian las neuronas en los humanos y, de hecho, en todos los animales. La señal tanto en las neuronas como en las hojas de Dionaea puede inhibirse mediante medicamentos que obstruyen los canales de iones que se abren en las membranas cuando la señal eléctrica atraviesa la célula. Cuando Volkov trató previamente sus plantas con una sustancia química que inhibe los canales de potasio en las neuronas humanas, por ejemplo, las trampas no se cerraron cuando se las tocaba ni al recibir las descargas eléctricas.[8]
MEMORIA DE LOS TRAUMAS A LARGO PLAZO
A mediados del siglo XX, el botánico checo Rudolf Dostál llevó a cabo unos siniestros estudios en plantas de lo que denominó «memoria morfogenética».[9] La memoria morfogenética es un tipo de memoria que influye posteriormente en la forma de la planta. En otras palabras: una planta puede experimentar un estímulo en un momento dado, como un desgarro en una hoja o una fractura de una rama, y en un inicio no resultar afectada, pero, cuando se produce un cambio de las condiciones ambientales, puede recordar esa experiencia pasada y reaccionar modificando su crecimiento.
Los experimentos de Dostál con plántulas de lino ilustran lo que entendía por memoria morfogenética. Para apreciar plenamente los experimentos de Dostál en este ámbito debemos entender un poco más la anatomía de una planta. Los plantones de lino brotan con dos grandes hojas llamadas «cotiledones». En el centro de esos dos cotiledones se encuentra lo que llamamos una «yema» o un «brote apical», que crece verticalmente a partir del tallo central de la planta. A medida que esta yema crece, emergen bajo ella dos brotes laterales a cada lado, cada uno de ellos encarado a una hoja. En condiciones normales, estos brotes laterales permanecen inactivos (no se desarrollan). Sin embargo, si la yema apical sufre algún daño o se corta, los brotes laterales empiezan a crecer y ampliarse y cada uno de ellos forma una nueva rama de la que cada brote lateral pasa a ser la yema apical. La represión de los brotes laterales por parte de la yema apical recibe el nombre de «dominio apical», y eliminarla constituye la base de la poda de las plantas. Cuando vemos a un jardinero podar los setos de delante de una casa, en realidad, si poda correctamente, lo que hace es cortar la yema apical de cada rama para alentar a que crezcan más brotes laterales y nuevas ramas.
Los robles, los pinos y sus hermanos en los bosques han visto tantas albas y crepúsculos, tantas estaciones pasar y tantas generaciones caer en el silencio que es fácil preguntarse qué nos contarían los árboles si tuvieran lenguas para hablar… o si nosotros tuviéramos oídos lo bastante agudos para entenderlos.
MAUD VAN BUREN,
Quotations for Special Occasions
7 ¿Qué recuerda una planta? (*)
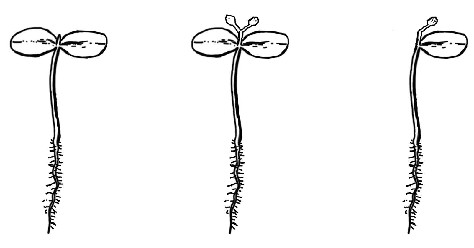
En esta ilustración vemos tres plántulas de lino (Linum usitatissimum). La imagen de la izquierda muestra un plantón de dos semanas de edad con dos cotiledones y una yema apical (el pequeño bultito entre las dos hojas). La imagen central muestra una plántula similar después de que se haya decapitado la yema apical y los dos brotes laterales se hayan desarrollado durante una semana aproximadamente. Y la imagen de la derecha muestra un plantón al que se cortó el cotiledón izquierdo antes de decapitar la yema apical.
En condiciones normales, si la yema apical se poda, ambos brotes laterales crecen de manera uniforme. Pero Dostál averiguó que, si eliminaba uno de los cotiledones antes de decapitar la yema apical, el único brote lateral que crecía era el que se encontraba cerca de la hoja restante.[10] Este resultado puede parecer un caso clásico de estímulo seguido de una reacción. Y ahora es cuando viene lo verdaderamente interesante. Cuando Dostál repitió el experimento e iluminó la planta con luz roja, el brote lateral más cercano al cotiledón ausente creció, hecho que reveló que cada brote lateral conserva el potencial de desarrollarse.
Michel Thellier, de la Universidad de Ruan, en el norte de Normandía, tomó el testigo de las investigaciones de Dostál. Thellier, integrante de la Academia de Ciencias de Francia, apreció que después de decapitar la yema apical de la planta de su elección, la Bidens pilosa (también conocida como amor seco), ambos brotes laterales empezaban a desarrollarse de manera más o menos uniforme.[11] En cambio, si se limitaba a dañar uno de los cotiledones solo crecía el brote lateral más cercano a la hoja sana. Thellier no tuvo que mutilar el cotiledón para obtener la reacción; le bastó con pinchar cuatro veces la hoja con una aguja en el momento de realizar la decapitación y aquella leve herida fue suficiente para estimular el crecimiento asimétrico de los brotes laterales.
En este caso, ¿dónde entra en acción la memoria de las plantas en lo que parece ser otro fenómeno clásico de estímulo y reacción? Pues bien, en ocasiones, durante sus experimentos, Thellier dilataba el tiempo que transcurría entre que dañaba la hoja y decapitaba la yema principal… hasta dos semanas. Y descubrió que el brote lateral más alejado del cotiledón dañado era el que crecía, mientras que el otro no lo hacía. Thellier sabía que, de alguna manera, el amor seco tenía que retener esa experiencia «traumática» y contar con un mecanismo para rememorarla una vez se cortaba la yema central, aunque ello sucediera muchos días después.
El siguiente experimento corroboró la idea de que el brote del amor seco recordaba cuál de sus hojas vecinas estaba dañada. En esta ocasión, Thellier clavó las agujas a uno de los cotiledones, tal como había hecho previamente, pero varios minutos después arrancó ambas hojas. Y descubrió que la planta retenía el recuerdo de los pinchazos: una vez cortada la yema central, el brote lateral opuesto al cotiledón dañado original crecía más que el situado en el lado de dicha hoja. El jurado aún está deliberando sobre cómo se almacena esta información en la yema central, pero una opción prometedora es que la señal esté conectada de algún modo con la auxina, la hormona de la cual hemos hablado en el capítulo 6.
EL GRAN ESCALOFRÍO
Trofim Denísovich Lysenko fue célebre por su repercusión en la ciencia en la Unión Soviética.[12] Rechazó la genética mendeliana clásica (basada en el principio de que todas las características son el resultado de genes heredados) y defendió la idea de que el entorno lleva al desarrollo de caracteres adaptativos (como la ceguera en los topos que viven en una oscuridad constante) que pueden transmitirse a generaciones posteriores. Esta teoría evolutiva, originalmente expuesta por el célebre naturalista francés Jean-Baptiste Lamarck en los albores del siglo XIX, encajaba a la perfección con la ideología dominante en la época en la que Lysenko llevó a cabo sus investigaciones, que sostenía que el proletariado podía modificarse mediante el entorno. La clase establecida soviética estaba tan encandilada con Lysenko que entre 1948 y 1964 fue ilegal en la Unión Soviética expresar cualquier disidencia de sus teorías. Política aparte, Lysenko realizó un descubrimiento fundamental en 1928 que a día de hoy sigue influyendo en la botánica.
Los agricultores soviéticos cultivan lo que se conoce como trigo de invierno, un trigo que se siembra en otoño, germina antes de las temperaturas gélidas invernales y permanece adormecido hasta que el suelo vuelve a calentarse en la primavera, momento en el cual florece. El trigo de invierno no florece y produce cereal en la primavera a menos que experimente un período de clima frío durante el invierno. Los últimos años de la década de 1920 fueron desastrosos para la agricultura soviética porque unos inviernos inusitadamente cálidos arrasaron con la mayoría de las plántulas de trigo de invierno, plántulas con las que los agricultores debían producir cereal para alimentar a millones de rusos.
Lysenko trabajó sin descanso para intentar conservar la poca cosecha que pudieran recolectar y encontrar un modo de garantizar que los inviernos cálidos no provocaran hambrunas en el futuro. Y descubrió que si introducía las semillas del trigo de invierno en un congelador antes de plantarlas podía inducir su germinación y florecimiento sin que hubieran vivido realmente un invierno prolongado. De este modo permitió a los agricultores plantar trigo en primavera y salvaguardó las cosechas de trigo de su país. Lysenko denominó este proceso «vernalización», palabra hoy asimilada como término general para cualquier tratamiento con frío, sea natural o artificial.
Pero Lysenko erró dramáticamente al afirmar que el rasgo del florecimiento inducido por el frío podía transmitirse a la generación siguiente. Estaba convencido de que su manipulación de las condiciones ambientales de las semillas del trigo provocaba un cambio permanente en la genética de la planta y, por supuesto, se equivocaba. Si bien la ciencia entiende hoy cómo el entorno influye en las características de una planta con el paso de las generaciones, tema que abordaremos enseguida, los esfuerzos de Lysenko de engranar su ideología política con su trabajo científico tuvieron consecuencias calamitosas para la genética en la Unión Soviética y limitaron seriamente la evolución de los estudios en la materia.[13]
Otros científicos sabían que algunas plantas necesitaban un clima frío para florecer (uno de los primeros informes surgió del Consejo de Agricultura de Ohio en 1857), pero Lysenko fue el primero en demostrar que este proceso podía manipularse artificialmente.[14] El cultivo de muchas plantas depende de las temperaturas frías del invierno; muchos árboles frutales solo florecen y dan fruto tras un frío invierno, y las semillas de lechuga y arabidopsis solo germinan tras una ola de frío. La ventaja económica de la vernalización es evidente: garantiza que tras el frío invernal una planta brotará o florecerá en primavera o verano y no durante otras épocas del año en las que la cantidad de luz y la temperatura podrían alentar su desarrollo.
Por ejemplo, los cerezos de Washington, D.C., suelen florecer por primera vez en el año en torno al 1 de abril, cuando hay unas doce horas de luz diurna. Pero en Washington también se disfrutan de doce horas de luz solar a mediados de septiembre y, sin embargo, esos mismos cerezos no florecen en otoño; de hacerlo, sus frutos no se desarrollarían nunca plenamente, porque se congelarían al aproximarse el invierno. Al florecer a principios de la primavera, los cerezos dan a sus frutos cinco meses completos para madurar. Y aunque la longitud del día es exactamente la misma en abril que en septiembre, los árboles son capaces de diferenciar entre ambos momentos. Saben que es abril porque recuerdan el invierno previo.
Hasta la pasada década no se había elucidado la base que explica que una plántula de trigo o un cerezo recuerden el invierno, y ello ha sido posible gracias a investigaciones realizadas con la arabidopsis, una planta de resultados contrastados. Las arabidopsis crecen en una amplia variedad de hábitats naturales, desde Noruega hasta las Islas Canarias. Las diferentes poblaciones de Arabidopsis thaliana reciben el nombre de «ecotipos». Los ecotipos de arabidopsis que crecen en los climas septentrionales precisan vernalización para florecer, mientras que no ocurre así con los que crecen en climas más cálidos. Esta necesidad de vernalización está codificada en los genes de los ecotipos del norte. Si se cruza una planta que precisa que haya invierno para florecer con una que no, la descendencia seguirá necesitando atravesar una ola de frío para crecer; genéticamente, la necesidad de frío es un rasgo dominante (tal como los ojos marrones son un rasgo dominante con respecto a los ojos azules en las personas). El gen específico implicado es el FLC, siglas en inglés de «flowering locus C». En su versión dominante, el FLC inhibe el florecimiento hasta que la planta ha experimentado un período de vernalización.
Una vez la planta atraviesa una fase de clima frío, el gen FLC deja de transcribirse, se desactiva. Pero ello no implica que las plantas empiecen a florecer de inmediato, sino solo que pueden florecer si confluyen otras condiciones, por ejemplo, relativas a la luz y la temperatura. De manera que la planta debe recordar de alguna manera que ha experimentado un clima frío para mantener desactivado el FLC por más que desde entonces las temperaturas hayan aumentado.
Muchos investigadores han intentado entender cómo la vernalización desactiva el FLC y lo mantiene desactivado. Estos estudios han subrayado que la epigenética está entrelazada en el recuerdo del invierno de una planta.[15] La epigenética alude a los cambios en la actividad de los genes que no requieren alteraciones en el código del ADN, tal como hacen las mutaciones, por más que tales cambios en la actividad genética se heredan de padres a hijos.[*] En muchos casos, la epigenética funciona mediante cambios en la estructura del ADN.
A nivel celular, el ADN se organiza en cromosomas, que son mucho más que simples cadenas de nucleótidos. La doble hélice del ADN se enrosca alrededor de unas proteínas denominadas «histonas» y forma lo que se conoce como cromatina. Esta cromatina puede enroscarse aún más, al igual que una banda elástica excesivamente enrollada, y compactar el ADN y las proteínas en estructuras muy condensadas y abarrotadas. Tales estructuras son dinámicas: distintas partes de la cromatina pueden desenrollarse o volverse a condensar. Los genes activos (los que se transcriben) están presentes en las partes de la cromatina que se han desenrollado, mientras que los inactivos se encuentran en las regiones más condensadas.[*]
Las proteínas histonas son uno de los factores clave que determinan la densidad con la que se empaca la cromatina y son fundamentales para entender cómo se activa el FLC. Los científicos han descubierto que el tratamiento con frío desencadena un cambio en la estructura de las histonas (un proceso llamado «metilación») que rodean el gen FLC, lo cual permite condensar mucho la cromatina. Esto desactiva el FLC y permite a la planta florecer. Este cambio epigenético (el tipo de histona alrededor del gen) se transmite de las células madre a las células hijas en las generaciones sucesivas y el gen FLC permanece inactivo en todas las células incluso después de concluido el período de clima frío. Una vez desactivado el gen FLC, las plantas pueden esperar a que el resto de las condiciones ambientales sean idóneas para florecer. En las plantas perennes como los robles y las azaleas, que florecen una vez al año, el gen FLC tiene que reactivarse después de que la planta haya dado flor y hasta después de transcurrido el invierno siguiente para inhibir un florecimiento promiscuo atemporal. Para ello, las células tienen que reprogramar el código de sus histonas, que abre la cromatina alrededor del gen FLC y lo reactiva.[16] En la actualidad se estudia cómo se produce este hecho y cómo se regula.[17]
Este y otros mecanismos epigenéticos explican que las plantas recuerden múltiples condiciones ambientales. Ahora bien, la memoria epigenética no es específica de las plantas y forma la base de muchos procesos biológicos y enfermedades. La epigenética ha propiciado un cambio de paradigma en la biología porque se opone al concepto genético clásico de que los únicos cambios que pueden transmitirse de célula a célula son los que están presentes en la secuencia del ADN. Lo verdaderamente asombroso acerca de la epigenética es que facilita el recuerdo no solo de estación en estación en un mismo organismo, sino de generación en generación.
EN CADA GENERACIÓN…
Los recuerdos se transmiten activamente de una generación a la siguiente mediante rituales, cuentos orales, etc. Sin embargo, la memoria transgeneracional que abarca la epigenética es completamente distinta. Este tipo de memoria suele implicar información acerca de estresores ambientales o físicos que se transmiten de padres a hijos. El laboratorio de Barbara Hohn en Basilea, Suiza, fue el primero en proporcionar pruebas de la existencia de esta memoria transgeneracional.[18] Hohn y sus colegas sabían que las condiciones que crean estrés en una planta, como la luz ultravioleta o un ataque de patógenos, provocan cambios en su genoma que derivan en nuevas combinaciones de ADN.
En términos ecológicos, estos cambios inducidos por el estrés tienen sentido porque, como cualquier otro organismo, una planta necesita hallar modos de sobrevivir bajo tensión, y una de las maneras que tiene de hacerlo es mediante nuevas variaciones genéticas. El fascinante estudio de Hohn demostró que las plantas estresadas no solo crean nuevas combinaciones de ADN, sino que sus vástagos también las crean aunque no se hayan visto directamente expuestos a ninguna tensión. El estrés experimentado por las progenitoras provocó un cambio heredable estable que se transmitió a sus retoños, los cuales se comportaban como si también hubieran vivido ese estrés. Recordaban que sus progenitoras lo habían padecido y reaccionaban de manera similar a ellas.
Usar la palabra «recordaban» en este contexto puede parecer poco ortodoxo, pero analicémoslo a la luz de las tres etapas de la memoria que hemos descrito al principio de este capítulo: los padres formaron el recuerdo del estrés, lo retuvieron y se lo transmitieron a sus vástagos, y dichos vástagos recuperaron la información y reaccionaron con acuerdo a ella, en este caso con nuevos cambios genómicos.
Las implicaciones de este estudio son amplias. Un estrés ambiental causa un cambio hereditario que se transmite a generaciones sucesivas. Ello encaja a la perfección con las teorías de Jean-Baptiste Lamarck, quien, como se recordará, afirmaba que la evolución se basaba en la herencia de caracteres adquiridos. Las plantas de Hohn, tras haber sido sometidas a un estrés con rayos ultravioleta o patógenos, adquirieron la característica de una variación genética incrementada y se la transmitieron a toda su progenie (¡y hay que tener en cuenta que una sola planta de arabidopsis produce miles de semillas!). Este hecho no puede explicarse por mutaciones en la secuencia del ADN de las plantas estresadas, porque, a lo sumo, en tal caso se transmitiría a un porcentaje muy reducido de los vástagos. Por otro lado, si el estrés indujo un cambio epigenético, este pudo tener lugar en todas las células de golpe, incluidas las del polen y los óvulos, y transferirse a toda la generación siguiente, así como a las futuras. Si bien los científicos siguen especulando acerca de la naturaleza del cambio epigenético involucrado en estos recuerdos, sigue siendo un enigma.
Igor Kovalchuk concibió un estudio de seguimiento en el que incluyó otros estresores de la variación genética en plantas y su progenie, incluidos el calor y la sal.[19] Demostró que estas distintas agresiones del medio ambiente aumentan la frecuencia de las reorganizaciones genómicas no solo en la generación de los progenitores sino también en la segunda generación. Los resultados obtenidos por Kovalchuk fueron fascinantes porque revelaron aún más que eso: no solo la segunda generación de plantas mostraba una variación genética aumentada, cosa que confirmaba los resultados de Hohn, sino que, además, era más tolerante a los diversos estresores. En otras palabras, los progenitores estresados generaron retoños que, en condiciones adversas, prosperaban mejor que las plantas normales. Casi con total seguridad, las distintas tensiones ambientales inducen cambios epigenéticos en la estructura de la cromatina de los progenitores, cambios que hereda su progenie. Creemos que así ocurre porque el equipo de Kovalchuk demostró que si trataban a los retoños con una sustancia química que borraba la información epigenética, esas mismas plantas perdían la capacidad de prosperar bajo el estrés ambiental. Los resultados de Hohn no contaron con la aceptación universal, como ocurre con muchos estudios científicos que provocan cambios de paradigma.[20] No obstante, un número creciente de ejemplos está cimentando la idea de la memoria transgeneracional. Por ejemplo, mi colega Georg Jander de la Cornell University demostró que «nietas» de plantas de arabidopsis atacadas por orugas seguían reaccionando a la defensiva generando ácido jasmónico en mayores cantidades, el cual provocaba que las orugas crecieran menos de un 50 por ciento de lo normal.[21] Esta memoria transgeneracional dependía de otro mecanismo epigenético en el que participaban pequeñas moléculas de ARN. Cada vez existe un mayor consenso con respecto a que estos resultados, como otros adicionales, anuncian una nueva era de la genética.[22] La idea de que el estrés genere recuerdos que se transmiten de generación en generación se sustenta en un número creciente de estudios realizados no solo en plantas, sino también en animales. En todos los casos, esta «memoria» se basa en alguna forma de herencia epigenética.
¿MEMORIA INTELIGENTE?
Es incuestionable que las plantas tienen la capacidad de retener y recuperar información biológica. Sin embargo, eso no significa que lo recuerden todo. De hecho, olvidan mucho más de lo que recuerdan, sobre todo en lo tocante a las tensiones.[23] Los recuerdos, que conducen a una respuesta determinada, podrían resultar útiles en un entorno con cambios predecibles y recurrentes. En cambio, en un entorno estable o impredecible, la mejor estrategia para una planta consiste en regresar a la situación previa al estrés o, lo que es lo mismo, en «olvidar» que dicho estrés se ha producido. Podemos entender esto a título personal planteándonos la siguiente pregunta: ¿de qué nos sirve recordar algo si no nos ayuda a actuar de manera distinta en el futuro? Estudios muy recientes señalan que el equilibrio entre recuerdo y olvido está influido por el período de recuperación tras la tensión, es decir, por el lapso transcurrido desde el estrés previo. Mecánicamente, este factor puede estar mediado por la estabilidad de determinadas moléculas de ARNm en la célula.
La intuición nos dice que los recuerdos de las plantas son muy distintos de los recuerdos detallados y teñidos de sentimientos que nosotros rememoramos a diario. Sin embargo, en un nivel básico, los comportamientos de distintas plantas descritos en este capítulo son tipos de memoria correctivos. La forma de enroscarse del zarcillo, el cierre de la venus atrapamoscas y el recuerdo de las tensiones ambientales en el caso de la arabidopsis engloban los procesos de formarse un recuerdo del evento, retenerlo durante períodos de tiempo claros y recordarlo en un momento posterior para obtener una respuesta específica que favorezca el desarrollo.[24]
Muchos de los mecanismos implicados en la memoria de las plantas también tienen una función en la memoria humana, incluidos la epigenética y los gradientes electroquímicos. Dichos gradientes son fundamentales para las conexiones neuronales de nuestros cerebros, la sede de la memoria tal como la mayoría la entendemos. En el transcurso de los últimos años, la botánica ha descubierto que las células vegetales no solo se comunican mediante corrientes eléctricas (tal como hemos visto en varios capítulos), sino que, además, las plantas contienen proteínas que en los humanos y otros animales funcionan como neurorreceptores. Un ejemplo perfecto es el receptor de glutamato. Los receptores de glutamato del cerebro son fundamentales para la comunicación neuronal, la formación de recuerdos y el aprendizaje, y existen una serie de medicamentos neuroactivos que los atacan. De ahí que a un equipo de científicos de la New York University les sorprendiera descubrir que las plantas contienen receptores de glutamato y que la arabidopsis es sensible a medicamentos neuroactivos que alteran la actividad de dichos receptores.[25]
¿De qué podían servirles a las plantas proteínas neurorreceptoras como el receptor de glutamato, si tenemos en cuenta que carecen de neuronas?[26] Estudios subsiguientes llevados a cabo por José Feijó y su equipo en Portugal demuestran que, en las plantas, estos receptores actúan en la señalización entre células de un modo muy similar a cómo se comunican entre sí las neuronas en los seres humanos. Y uno no puede más que maravillarse ante la función evolutiva de los «receptores cerebrales» en las plantas. Quizá las similitudes entre la función cerebral humana y la fisiología vegetal sean mayores de lo que suponíamos.
Los recuerdos de las plantas, como la memoria inmunológica de los humanos, no responden a la memoria semántica o episódica, según la definición de Tulving, sino más bien a la memoria procedimental, al recuerdo sobre cómo hacer las cosas, y tales recuerdos dependen de la capacidad de notar estímulos externos. Tulving planteó que cada uno de los tres niveles de memoria está relacionado con un nivel creciente de conciencia.[27] La memoria procedimental va asociada a la conciencia anoética, la memoria semántica a la conciencia noética y la memoria episódica a la conciencia autonoética. Es evidente que las plantas no encajan en la definición de la conciencia asociada con la memoria semántica o episódica. Pero, tal como recogía un artículo de opinión reciente: «El nivel inferior de conciencia característico de la memoria procedimental, la conciencia anoética, alude a la capacidad de los organismos de percibir estímulos externos e internos y reaccionar a ellos, cosa de la que todas las plantas y animales simples son capaces».[28] Y ello nos lleva a la pregunta más enigmática de todas: si las plantas presentan distintos tipos de memoria y tienen alguna forma de conciencia, ¿habría que considerarlas inteligentes?
(*) Sacado de: Chamovitz, D. (2012). Lo que las plantas saben (1.ª ed., p. cap. 7 ). USA: Scientific American / Farrar, Straus and Giroux .